Plants are the source of our food, medicine, construction materials, and the foundation of ecosystems. How can we improve the productivity and resilience of plants? What knowledge, technologies, and tools do we need to generate? These are the long-term questions Zhiyong Wang's lab aim to answer in their research.
Plant growth and survival depend on cellular signaling mechanisms through which plant cells monitor and respond to hormonal signals, environmental cues, and internal nutrient status. Brassinosteroid (BR) is a major growth-promoting hormone that effects on plant height, size, and biomass accumulation.
Plant growth and development are also highly sensitive to environmental signals such as light/dark, temperature, and pathogens. Of course, plant growth depends on nutrients including nitrogen and sugars (product of photosynthesis), and nutrient-sensing mechanisms, such as the Target of Rapamycin (TOR) kinase or O-glycosyltransferases (SPINDLY and SECRET AGENT), are essential for viability.
Zhiyong Wang's research dissects the molecular mechanisms underlying growth responses to these internal and external factors, which have major impacts on plant growth and resilience. To achieve a comprehensive and mechanistic understanding of the growth regulatory system, his lab uses broad research approaches and technologies, including genomics, proteomics, chemical proteomics, microscopy, computation, and structural biology.
Accomplishments
The Wang Lab's research has established the framework of molecular networks that explain how nutritional, hormonal, and environmental signals coordinate the cellular decisions of growth, immunity, and acclimation. Most of the former postdocs and students who made these important discoveries are now leading their own labs in academic institutions.
Among the major achievements of our research, Zhiyong Wang's team has illustrated:
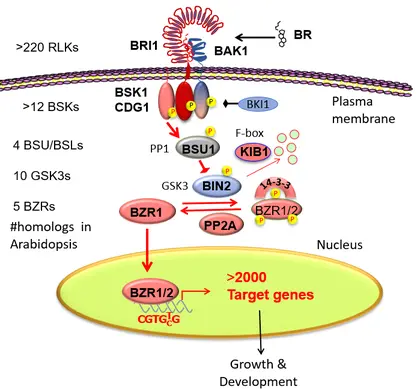
The full brassinosteroid (BR) signaling pathway from the receptor kinase BRI1 to nuclear transcription factor BZR1 and its thousands of target genes.
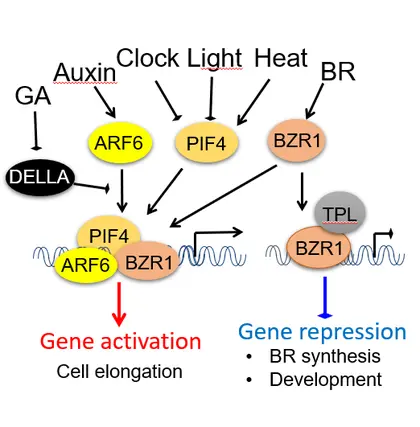
The growth co-regulation by key growth hormones (BR, auxin, gibberellin) and environmental signals (light and temperature) through direct interactions among their responsive transcription factors, a signal integration mechanism named BAP/D module
The spatiotemporal actions of BR in patterning growth and development in the shoot and root tips.
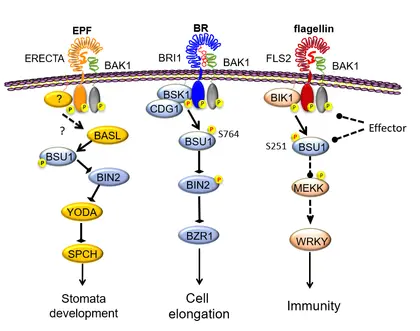
The mechanisms of crosstalk and component-sharing between BR/BRI1 and other receptor kinase pathways that regulate stomata development and immunity.
The expansive BR-response phosphorylation network controlled by the BIN2/GSK3 kinase.
The genetic variations in the BR-response cis-elements contribute to traits in maize.
The expansive nutrient-signaling networks of protein posttranslational modifications by O-linked β-N-acetylglucosamine (O-GlcNAc) and O-fucose.
The significant overlaps between the BR-regulated phosphorylation network and the nutrient-dependent O-glycosylation networks.
Ongoing Work:
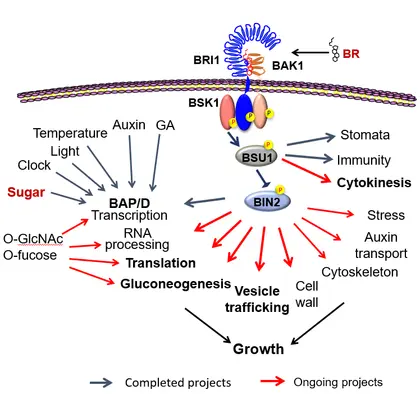
Building upon a large amount of solid data and converging discoveries while taking advantage of the in-house mass spectrometry facility/technologies, our current research continues to make exciting progress toward answering important scientific questions. These include:
How does BR-dependent phosphorylation regulate membrane trafficking, an essential aspect of cell growth?
How do the BR-signaling proteins regulate cytokinesis in plants?
How do cells maintain cell wall integrity during hormone-induced cell expansion?
How do O-GlcNAcylation and O-fucosylation mediate sugar regulation of protein functions and cellular/developmental/physiological processes?
How do BR and sugar signaling, through phosphorylation and O-glycosylation, respectively, co-regulate metabolism and growth?
How do phosphorylation and O-glycosylation crosstalk on common target proteins?
These projects are led by individual postdocs and graduate students, who collaborate and support each other, under my guidance. Together, we are advance a systems-level mechanistic understanding of plant growth and acclimation, and we identify targets and strategies for improving plant productivity and resilience.
Looking Ahead:
What are the main challenges that we still need to overcome? What are the opportunities provided by accumulating knowledge and advancing technologies?
We need to develop tools that enable spatiotemporal manipulation of specific signaling events, and we are developing such tools using nanobodies, molecular sensors, and chemicals/drugs. We would like to expand our research into non-model plants of economic or ecological importance. To do this, we need funding and people to replicate in crops (e.g. maize) some of the productive proteomic experiments (e.g. proximity labeling and O-glycosylation profiling) that we have done in Arabidopsis. We also need to develop better transformation methods to easily transform plants that are difficult or impossible to transform with current methods, and we are testing some novel ideas.
The rapid development in technologies presents exciting opportunities for life science. For example, structures of nearly all proteins can now be predicted by AlphaFold and visualized by cryoEM. This makes it possible to carry out structure-based drug discovery for plant biology. We are using combinations of virtual and experimental screening approaches to identify chemical inhibitors and modulators of plant proteins, developing chemical tools useful for basic research and agricultural application.
Contact Zhiyong Wang
The Wang Lab strives to push the frontier and hope to one day make a real positive impact on our world. If you are interested in joining us, or supporting us, reach out via email to zwang@carnegiescience.edu.
Plants often adapt to adverse or stress conditions via differential growth. The trans-Golgi network (TGN) has been implicated in stress responses, but it is not clear in what capacity it mediates adaptive growth decisions. In this study, we assess the role of the TGN in stress responses by exploring the previously identified interactome of the Transport Protein Particle II (TRAPPII) complex required for TGN structure and function. We identified physical and genetic interactions between AtTRAPPII and shaggy-like kinases (GSK3/AtSKs) and provided in vitro and in vivo evidence that the TRAPPII phosphostatus mediates adaptive responses to abiotic cues. AtSKs are multifunctional kinases that integrate a broad range of signals. Similarly, the AtTRAPPII interactome is vast and considerably enriched in signaling components. An AtSK-TRAPPII interaction would integrate all levels of cellular organization and instruct the TGN, a central and highly discriminate cellular hub, as to how to mobilize and allocate resources to optimize growth and survival under limiting or adverse conditions.
Brassinosteroid (BR) signaling leads to the nuclear accumulation of the BRASSINAZOLE-RESISTANT 1 (BZR1) transcription factor, which plays dual roles in activating or repressing the expression of thousands of genes. BZR1 represses gene expression by recruiting histone deacetylases, but how it activates transcription of BR-induced genes remains unclear. Here, we show that BR reshapes the genome-wide chromatin accessibility landscape, increasing the accessibility of BR-induced genes and reducing the accessibility of BR-repressed genes in Arabidopsis. BZR1 physically interacts with the BRAHMA-associated SWI/SNF (BAS)-chromatin-remodeling complex on the genome and selectively recruits the BAS complex to BR-activated genes. Depletion of BAS abrogates the capacities of BZR1 to increase chromatin accessibility, activate gene expression, and promote cell elongation without affecting BZR1's ability to reduce chromatin accessibility and expression of BR-repressed genes. Together, these data identify that BZR1 recruits the BAS complex to open chromatin and to mediate BR-induced transcriptional activation of growth-promoting genes.
By directly altering microscopic interactions, pressure provides a powerful tuning knob for the exploration of condensed phases and geophysical phenomena1. The megabar regime represents an interesting frontier, in which recent discoveries include high-temperature superconductors, as well as structural and valence phase transitions2-6. However, at such high pressures, many conventional measurement techniques fail. Here we demonstrate the ability to perform local magnetometry inside a diamond anvil cell with sub-micron spatial resolution at megabar pressures. Our approach uses a shallow layer of nitrogen-vacancy colour centres implanted directly within the anvil7-9; crucially, we choose a crystal cut compatible with the intrinsic symmetries of the nitrogen-vacancy centre to enable functionality at megabar pressures. We apply our technique to characterize a recently discovered hydride superconductor, CeH9 (ref.10). By performing simultaneous magnetometry and electrical transport measurements, we observe the dual signatures of superconductivity: diamagnetism characteristic of the Meissner effect and a sharp drop of the resistance to near zero. By locally mapping both the diamagnetic response and flux trapping, we directly image the geometry of superconducting regions, showing marked inhomogeneities at the micron scale. Our work brings quantum sensing to the megabar frontier and enables the closed-loop optimization of superhydride materials synthesis.
Wind is one of the most prevalent environmental forces entraining plants to develop various mechano-responses, collectively called thigmomorphogenesis. Largely unknown is how plants transduce these versatile wind force signals downstream to nuclear events and to the development of thigmomorphogenic phenotype or anemotropic response. To identify molecular components at the early steps of the wind force signaling, two mechanical signaling-related phosphoproteins, identified from our previous phosphoproteomic study of Arabidopsis touch response, mitogen-activated protein kinase kinase 1 (MKK1) and 2 (MKK2), were selected for performing in planta TurboID (ID)-based quantitative proximity-labeling (PL) proteomics. This quantitative biotinylproteomics was separately performed on MKK1-ID and MKK2-ID transgenic plants, respectively, using the genetically engineered TurboID biotin ligase expression transgenics as a universal control. This unique PTM proteomics successfully identified 11 and 71 MKK1 and MKK2 putative interactors, respectively. Biotin occupancy ratio (BOR) was found to be an alternative parameter to measure the extent of proximity and specificity between the proximal target proteins and the bait fusion protein. Bioinformatics analysis of these biotinylprotein data also found that TurboID biotin ligase favorably labels the loop region of target proteins. A WInd-Related Kinase 1 (WIRK1), previously known as rapidly accelerated fibrosarcoma (Raf)-like kinase 36 (RAF36), was found to be a putative common interactor for both MKK1 and MKK2 and preferentially interacts with MKK2. Further molecular biology studies of the Arabidopsis RAF36 kinase found that it plays a role in wind regulation of the touch-responsive TCH3 and CML38 gene expression and the phosphorylation of a touch-regulated PATL3 phosphoprotein. Measurement of leaf morphology and shoot gravitropic response of wirk1 (raf36) mutant revealed that the WIRK1 gene is involved in both wind-triggered rosette thigmomorphogenesis and gravitropism of Arabidopsis stems, suggesting that the WIRK1 (RAF36) protein probably functioning upstream of both MKK1 and MKK2 and that it may serve as the crosstalk point among multiple mechano-signal transduction pathways mediating both wind mechano-response and gravitropism.
By directly altering microscopic interactions, pressure provides a powerful tuning knob for the exploration of condensed phases and geophysical phenomena1. The megabar regime represents an interesting frontier, in which recent discoveries include high-temperature superconductors, as well as structural and valence phase transitions2-6. However, at such high pressures, many conventional measurement techniques fail. Here we demonstrate the ability to perform local magnetometry inside a diamond anvil cell with sub-micron spatial resolution at megabar pressures. Our approach uses a shallow layer of nitrogen-vacancy colour centres implanted directly within the anvil7-9; crucially, we choose a crystal cut compatible with the intrinsic symmetries of the nitrogen-vacancy centre to enable functionality at megabar pressures. We apply our technique to characterize a recently discovered hydride superconductor, CeH9 (ref. 10). By performing simultaneous magnetometry and electrical transport measurements, we observe the dual signatures of superconductivity: diamagnetism characteristic of the Meissner effect and a sharp drop of the resistance to near zero. By locally mapping both the diamagnetic response and flux trapping, we directly image the geometry of superconducting regions, showing marked inhomogeneities at the micron scale. Our work brings quantum sensing to the megabar frontier and enables the closed-loop optimization of superhydride materials synthesis.
Protein O-glycosylation is a nutrient signaling mechanism that plays an essential role in maintaining cellular homeostasis across different species. In plants, SPINDLY (SPY) and SECRET AGENT (SEC) posttranslationally modify hundreds of intracellular proteins with O-fucose and O-linked N-acetylglucosamine, respectively. SPY and SEC play overlapping roles in cellular regulation, and loss of both SPY and SEC causes embryo lethality in Arabidopsis (Arabidopsis thaliana). Using structure-based virtual screening of chemical libraries followed by in vitro and in planta assays, we identified a SPY O-fucosyltransferase inhibitor (SOFTI). Computational analyses predicted that SOFTI binds to the GDP-fucose-binding pocket of SPY and competitively inhibits GDP-fucose binding. In vitro assays confirmed that SOFTI interacts with SPY and inhibits its O-fucosyltransferase activity. Docking analysis identified additional SOFTI analogs that showed stronger inhibitory activities. SOFTI treatment of Arabidopsis seedlings decreased protein O-fucosylation and elicited phenotypes similar to the spy mutants, including early seed germination, increased root hair density, and defective sugar-dependent growth. In contrast, SOFTI did not visibly affect the spy mutant. Similarly, SOFTI inhibited the sugar-dependent growth of tomato (Solanum lycopersicum) seedlings. These results demonstrate that SOFTI is a specific SPY O-fucosyltransferase inhibitor that can be used as a chemical tool for functional studies of O-fucosylation and potentially for agricultural management.